Что содержит рнк. Строение рнк. Структура молекулы рибнуклеиновой кислоты
Для поддержания жизни в живом организме происходит множество процессов. Некоторые из них мы можем наблюдать - дыхание, прием пищи, избавление от продуктов жизнедеятельности, получение информации органами чувств и забывание этой информации. Но большая часть химических процессов скрыта от глаз.
Справка. Классификация
По-научному, обмен веществ это метаболизм.
Метаболизм обычно делят на две стадии:в ходе катаболизма сложные органические молекулы распадаются на более простые, с получением энергии; (энергия тратится)
в процессах анаболизма затрачивается энергия на синтез из простых молекул сложных биомолекул. (энергия запасается)
Биомолекулы, как видно выше, делятся на малые молекулы и большие.
Малые:Липиды (жиры), фосфолипиды, гликолипиды, стеролы, глицеролипиды,
Витамины
Гормоны, нейромедиаторы
Метаболиты
Большие:
Мономеры, олигомеры и полимеры.
Мономеры Олигомеры Биополимеры
Аминокислоты Олигопептиды Полипептиды, белкиМоносахариды Олигосахариды Полисахариды (крахмал, целлюлоза)
Нуклеотиды Олигонуклеотиды Полинуклеотиды, (ДНК, РНК)
В столбце биополимеры находятся полинуклеотиды. Именно здесь находится рибонуклеиновая кислота - объект статьи.
Рибонуклеиновые кислоты. Строение, назначение.

На рисунке показана молекула РНК.
Нуклеиновые кислоты ДНК и РНК присутствуют в клетках всех живых организмов и выполняют функции по хранению, передаче и реализации наследственной информации.
Сходство и отличие РНК и ДНК
Как видно, есть внешнее сходство с известной структурой молекулы ДНК (дезоксирибонуклеиновой кислотой).Однако, РНК может иметь как двухцепочечную структуру, так и одноцепочечную.
Нуклеотиды (пяти- и шестиугольники на рисунке)
Кроме того, нить РНК состоит из четырех нуклеотидов (или азотистых оснований, что одно и то же): аденин, урацил, гуанин и цитозин.
Нить ДНК же состоит из другого набора нуклеотидов: аденин, гуанин, тимин и цитозин.
Химическое строение полинуклеотида РНК:

Как видим, имеются характерные нуклеотиды урацил (для РНК) и тимин (для ДНК).
Все 5 нуклеотидов на рисунке:

Шестиугольники на рисунках - это бензольные кольца, в которые, вместо углерода, встраиваются другие элементы, в данном случае, это азот.
Бензол. Для справки.
Химическая формула бензола - C6H6. Т.е. в каждом угле шестиугольника находится атом углерода. 3 дополнительные внутренние линии в шестиугольнике указывают на наличие двойных ковалентных связей между этими атомами углерода. Углерод - элемент 4 группы периодической таблицы Менделеева, следовательно, у него 4 электрона могут образовать ковалентную связь. На рисунке - одна связь - с электроном водорода, вторая - с электроном углерода слева и еще 2 - с 2 электронами углерода справа. Впрочем, физически существует единое электронное облако, охватывающее все 6 атомов углерода бензола.
Соединение азотистых оснований
Комплементарные нуклеотиды друг с другом сцепляются (гибридизуются) с помощью водородных связей. Аденин комплементарен урацилу, а гуанин - цитозину. Чем длиннее на данной РНК комплементарные участки, тем прочнее будет образуемая ими структура; наоборот, короткие участки будут нестабильными. Это определяет функцию конкретной РНК.На рисунке фрагмент комплементарного участка РНК. Азотистые основания закрашены синим цветом

Структура РНК
Сцепление многих групп нуклеотидов образуют РНК-шпильки (первичная структура):
Множество шпилек в ленте сцепляются в двойную спираль. В развернутом виде такая структура напоминают дерево (Вторичная структура):

Спирали так же взаимодействуют друг с другом (третичная структура). Видно, как разные спирали соединены друг с другом:

Другие РНК сворачиваются аналогично. Напоминает набор лент (четвертичная структура).

Заключение
Для вычисления конформаций, которые примут РНК, по их первичной последовательности существуютИ урацил (в отличие от ДНК, содержащий вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусов.
Основные функции РНК в клеточных организмах - это шаблон для трансляции генетической информации в белки и поставка соответствующих аминокислот к рибосомам. В вирусах является носителем генетической информации (кодирует белки оболочки и ферменты вирусов). Вироиды состоят из кольцевой молекулы РНК и не содержат в себе других молекул. Существует гипотеза мира РНК , согласно которой, РНК возникли перед белками и были первыми формами жизни.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами - РНК-полимеразы. Затем матричные РНК (мРНК) участвуют в процессе, называемом трансляцией. Трансляция - это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечной РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а матричные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так мРНК участвуют в эукариотических матричных РНК и других процессах.
Кроме того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Ряда вирусов состоят из РНК, то есть у них она играет роль, которую в высших организмах выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК - первая молекула, способная к самовоспроизведению в добиологических системах.
История изучения РНК
Нуклеиновые кислоты были открыты в 1868 году швейцарским ученым Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus). Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты.
Значение РНК в синтезе белков было предположено в 1939 году в работе Торберна Оскара Касперссона, Жана Брачета и Джека Шульца. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при ее введении в ооциты образуется тот же самый белок.
В Советском Союзе в 1956-57 годах проводились работы (А. Белозерский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляют рибосомные РНК.
В 1959 году Северо Очоа получил Нобелевскую премию по медицине за открытие механизма синтеза РНК. Последовательность из 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холле, за что в 1968 году он получил Нобелевскую премию по медицине.
В 1967 Карл Везе предположил, что РНК имеют каталитические свойства. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служили и как молекулы хранения информации (сейчас эта роль выполняется ДНК) и как молекулы, которые катализировали метаболические реакции (сейчас это делают ферменты).
В 1976 Уолтер Фаерс и его группа из Гентского университета (Голландия) впервые определили последовательность генома РНК - содержащегося в вирусе, бактериофага MS2.
В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения. Примерно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микро-РНК, играют регуляторную роль в онтогенезе круглых червей.
Гипотеза о значении РНК в синтезе белков была высказана Торбьерном Касперссоном (Torbjörn Caspersson) на основе исследований 1937-1939 гг ., в результате которых было показано, что клетки, активно синтезирующие белок, содержат большое количество РНК. Подтверждение гипотезы было получено Юбером Шантренном (Hubert Chantrenne).
Особенности строения РНК
Нуклеотиды РНК состоят из сахара - рибозы, к которой в положении 1 "присоединена одна из основ: аденин, гуанин, цитозин или урацил. Фосфатная группа объединяет рибозы в цепочку, образуя связи с 3 "атомом углерода одной рибозы и в 5" положении другого. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК - можно назвать полианионом .
РНК транскрибируется как полимер четырех оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C)), но в «зрелой» РНК есть много модифицированных оснований и сахаров. Всего в РНК насчитывается около 100 различных видов модифицированных нуклеозидов, из которых:
- 2"-О-метилрибоза
наиболее частая модификация сахара;
- Псевдоуридин
- наиболее часто модифицированная основа, которая встречается чаще всего. В псевдоуридине (Ψ) связь между урацилом и рибозой не C - N, а C - C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК.
Еще одной модифицированной основой, о которой стоит сказать является - гипоксантин, деаминованний гуанин, нуклеозид которого носит название инозин . Инозин играет важную роль в обеспечении вырожденности генетического кода.
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК много пост-транскрипционных модификаций находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующих в образовании пептидной связи. Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырех нуклеотидов, в которой есть пара оснований аденин - гуанин.
Важная структурная особенность РНК, отличающая ее от ДНК - наличие гидроксильной группы в 2 "положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, которая наиболее часто наблюдается в ДНК. В А-форме глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2 "гидроксильной группы состоит в том, что конформационно пластичные, то есть, не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять.
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуется посредством водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры - стебель-петли, петли и псевдоузлы. В силу большого количества возможных вариантов спаривания оснований, предсказания вторичной структуры РНК - гораздо более сложная задача, чем структуры белков, но в настоящее время есть эффективные программы, например, mfold.
Примером зависимости функций молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES - структура на 5 "конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требует наличия особого модифицированного основания (кэпа) на 5" конце и белковых факторов инициации. Сначала IRES были обнаружены в вирусных РНК, но сейчас накапливается все больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса. Многие типы РНК, например, рРНК и мяРНК (мяРНК) в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у ) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами .
Матричная рибонуклеиновая кислота (мРНК, синоним - информационная РНК, иРНК)
- РНК, отвечающая за перенос информации о первичной структуре белков от ДНК к местам синтеза белков. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется при трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) .
Длина типичной зрелой мРНК составляет от нескольких сотен до нескольких тысяч нуклеотидов. Самые длинные мРНК отмечены у (+) оц РНК-содержащих вирусов, например пикорнавирусов, однако следует помнить, что у этих вирусов мРНК образует весь их геном.
Подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибировать из отдельных генов (например, рибосомальные РНК) или быть производными интронов. Классические, хорошо изученные типы некодирующих РНК - это транспортные РНК (тРНК) и рРНК, участвующие в процессе трансляции. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции - энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Транспортные (тРНК) - малые, состоящие из примерно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты к месту синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодону мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединенной к тРНК.
Рибосомальные РНК (рРНК) - каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырех типов рРНК синтезируются на полисомах. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеины, называемые рибосомами. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80% РНК, обнаруживается в цитоплазме эукариотической клетки.
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию.
Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и влияют через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводит к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградирует. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.
Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
1 . ДНК содержит сахар дезоксирибозу, РНК - рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
2.
Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил - неметилированная форма тимина.
3.
ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, намного короче и преимущественно одноцепочечные. Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образуют нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептид-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК.
Особенности функций:
1. Процессинг
Многие РНК принимают участие в модификации других РНК. Интроны вырезают из про-мРНК сплайсосомы, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированная в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышке и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связывается с РНК-мишенью путем образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК.
2. Трансляция
ТРНК присоединяют определенные аминокислоты в цитоплазме и направляется к месту синтеза белка на иРНК где связывается с кодоном и отдает аминокислоту которая используется для синтеза белка.
3. Информационная функция
У некоторых вирусов РНК выполняет те функции которые ДНК выполняет у эукариот. Также информационную функцию выполняет иРНК которая переносит информацию о белках и является местом его синтеза.
4. Регуляция генов
Некоторые типы РНК участвуют в регуляции генов увеличивая или уменьшая его активность. Это так называемые миРНК (малые интерферирующие РНК) и микро-РНК.
5. Каталитическая функция
Есть так называемые ферменты которые относятся к РНК они называются рибозимы. Эти ферменты выполняют различные функции и имеют своеобразное строение
сокр., RNA ) — линейный полимер, образованный ковалентно связанными рибонуклеотидными мономерами.Описание
Рибонуклеиновые кислоты (РНК) - полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания - аденин, цитозин, гуанин и урацил (в отличие от , содержащей вместо урацила тимин). Эти молекулы содержатся в всех живых организмов, а также в некоторых вирусах. У некоторых РНК служит носителем генетической информации. РНК, как правило, построены из одной полинуклеотидной цепи. Известны редкие примеры двухспиральных молекул РНК. Различают 3 основных типа РНК: рибосомную (рРНК), транспортную (тРНК) и информационную или матричную (иРНК, мРНК). Матричная РНК служит для передачи информации, закодированной в ДНК, рибосомам, синтезирующим . Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка. Однако подавляющее большинство разновидностей РНК не кодирует белок (например, тРНК и рРНК). Существуют и другие некодирующие РНК, например, РНК, ответственные за регуляцию генов и процессинг мРНК; РНК, катализирующие разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции, - энзимами, каталитические молекулы РНК называются рибозимами. Микро-РНК (размером 20–22 нуклеотидных пар) и малые интерферирующие РНК (миРНК, размером 20–25 нуклеотидных пар) способны уменьшать или увеличивать экспрессию генов через механизм РНК-интерференции. Специфические белки системы направляются при помощи микро- и миРНК к целевым последовательностям Мрнк и разрезают их, вследствие чего нарушается процесс трансляции. На основе механизма РНК-интерференции разработана перспективная новая технология рака, направленная на «выключение» (сайленсинг, от англ. silence - молчание) генов, отвечающих за рост и деление раковых клеток. В настоящее время активно разрабатываются методы доставки с помощью специализированных целевых миРНК в опухолевые клетки.
Авторы
- Народицкий Борис Савельевич
- Ширинский Владимир Павлович
- Нестеренко Людмила Николаевна
Источники
- Alberts B., Johnson A., Lewis J. et al. Molecular Biology of the Cell. 4th ed. - N.Y.: Garland Publishing, 2002. - 265 p.
- Рис Э., Стернберг М. Введение в молкулярную биологию. От клеток к атомам. - М.: Мир, 2002. - 154 с.
- Рибонуклеиновые кислоты // Википедия, свободная энциклопедия. - http://ru.wikipedia.org/wiki/Рибонуклеиновые_кислоты (дата обращения: 02.10.2009).
Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
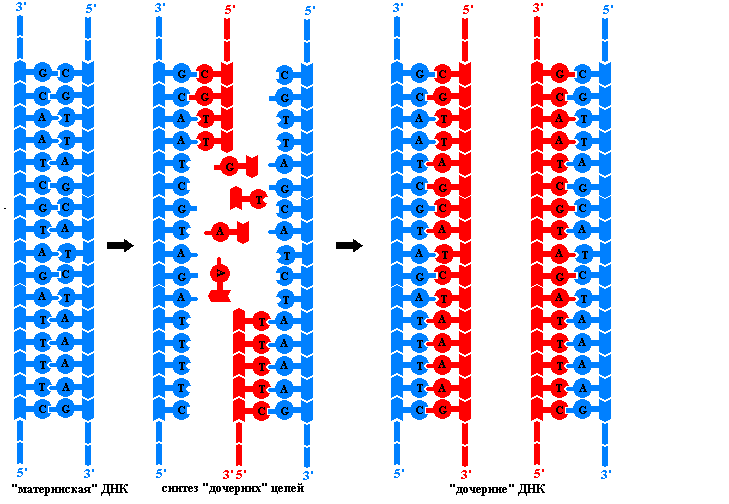
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.
Функции РНК различаются в зависимости от вида рибонуклеиновый кислоты.
1) Информационная РНК (и-РНК).
2) Рибосомная РНК (р-РНК).
3) Транспортная РНК (т-РНК).
4) Минорные (малые) РНК. Это молекулы РНК, чаще всего с небольшой молекулярной массой, располагающиеся в различных участках клетки (мембране, цитоплазме, органеллах, ядре и т.д.). Их роль до конца не изучена. Доказано, что они могут помогать созреванию рибосомной РНК, участвуют в переносе белков через мембрану клетки, способствуют редупликации молекул ДНК и т.д.
5) Рибозимы. Недавно выявленный вид РНК, принимающие активное участие в ферментативных процессах клетки в качестве фермента (катализатора).
6) Вирусные РНК. Любой вирус может содержать только один вид нуклеиновой кислоты: либо ДНК либо РНК. Соответственно, вирусы, имеющие в своём составе молекулу РНК, получили название РНК-содержащие. При попадании в клетку вируса данного типа может происходить процесс обратной транскрипции (образование новых ДНК на базе РНК), и уже вновь образовавшаяся ДНК вируса встраивается в геном клетки и обеспечивает существование, а также размножение возбудителя. Вторым вариантом сценария является образование комплиментарной РНК на матрице поступившей вирусной РНК. В этом случае, образование новых вирусных белков, жизнедеятельность и размножение вируса происходит без участия дезоксирибонуклеиновой кислоты только на основании генетической информации, записанной на вирусной-РНК. Рибонуклеиновые кислоты. РНК, строение, структуры, виды, роль. Генетический код. Механизмы передачи генетической информации. Репликация. Транскрипция
Рибосомная РНК.
На долю рРНК приходится 90% всей РНК клетки, она характеризуется метаболической стабильностью. У прокариот различают три различных типа рРНК с коэффициентами седиментации 23S,16S и 5S; у эукариот четыре типа:-28S, 18S,5S и 5,8S.
РНК этого типа локализованы в рибосомах и участвуют в специфическом взаимодействии с рибосомными белками.
Рибосомные РНК имеют форму вторичной структуры в виде которых двуспиральных участков, соединенных изогнутой одиночной цепью. Белки рибосомы связаны преимущественно с однотяжевыми участками молекулы.
Для рРНК характерно наличие модифицированных оснований, однако в значительно меньшем количестве, чем в тРНК. В рРНК присутствуют главным образом метилизированные нуклеотиды, причем метильные группы присоединены либо к основанию, либо к 2 / - OH- группе рибозы.
Транспортная РНК.
Молекулы тРНК представляют собой единую цепь, состоящую из 70-90 нуклеотидов, с молекулярной массой 23000-28000 и константой седиментации 4S. В клеточной РНК транспортная РНК составляет 10-20%. Молекулы тРНК обладают способностью ковалентно связываться с определенной аминокислотой и соединяться через систему водородных связей с одним из нуклеотидных триплетов молекулы мРНК. Таким образом, тРНК реализуют кодовое соответствие между аминокислотой и отвечающим ей кодоном мРНК. Для выполнения адапторной функции тРНК должны иметь вполне определенную вторичную и третичную структуру.
Каждая молекула тРНК обладает постоянной вторичной структурой, имеет форму двумерного клеверного листа и состоит из спиральных участков, образованных нуклеотидами одной и той же цепи, и расположенных между ними одноцепочечных петель. Количество спиральных областей достигает половины молекулы.Неспаренные последовательности образуют характерные структурные элементы (ветви),имеющие типичные ветви:
А) акцепторный стебель, на 3 / -OH конце которого в большинстве случаев расположен триплет ЦЦА. К карбоксильной группе концевого аденозина с помощью специфического фермента присоединяется соответствующая аминокислота;
Б) псевдоуридиновая или Т Ц-петля, состоит из семи нуклеотидов с обязательной последовательностью 5 / -Т ЦГ-3 / , в которой содержится псевдоуридин; предполагается что Т Ц-петля используется для связывания тРНК с рибосомой;
В) дополнительная петля-различная по размеру и составу в разных тРНК;
Г) антикодоновая петля состоит из семи нуклеотидов и содержит группу из трех оснований (антикодон), которая комплементарна триплету (кодону) в молекуле иРНК;
Д) дигидроуридиловая петля (D-петля), состоящая из 8-12 нуклеотидов и содержащая от одного до четырех дигидроуридиловых остатков;считается, что D-петля используется для связывания тРНК со специфическим ферментом (аминоацил-тРНК-синтетаза).
Третичная укладка молекул тРНК является весьма компактной и имеет Г-образную форму. Угол подобной структуры образован дигидроуридиновым остатком и Т Ц-петлей, длинное колено образует акцепторный стебель и Т Ц-петля, а короткое-D-петля и антикодоновая петля.
В стабилизации третичной структуры тРНК участвуют поливалентные катионы (Mg 2+ , полиамины), а также водородные связи между основаниями и фосфодиэфирным остовом.
Сложная постранственная укладка молекулы тРНК обусловлена множественными высокоспецифичными взаимодействиями как с белками, так и с другими нуклеиновыми кислотами (рРНК).
Транспортная РНК отличается от других типов РНК высоким содержанием минорных оснований-в среднем 10-12 оснований на молекулу, однако общее число их а тРНК растет по мере продважения организмов по эволюционной лестнице. В тРНК выявлены различные метилированные пуриновые (аденин, гуанин) и пиримидиновые (5-метилцитозин и рибозилтимин) основания, серосодержащие основания (6-тиоурацил), но наиболее распростран(6-тиоурацил), но наиболее распространенным минорным компонентом является псевдоуридин. Роль необычных нуклеотидов в молекулах тРНК пока не ясна, однако пологают, что чем ниже уровень митилирования тРНК, тем она менее активна и специфична.
Локализация модифицированных нуклеотидов строго фиксирована. Наличие минорных оснований в составе тРНК обуславливает устойчивасть молекул к действию нуклеаз и, кроме того, они участвуют в поддержании определенной структуры, так как подобные основания не способны к нормальному спариванию и препятствуют образованию двойной спирали. Таким образом, наличие модифицированных оснований в составе тРНК обуславливает не только её структуру, но также и многие специальные функции молекулы тРНК.
В большинстве клеток эукариот содержится набор различных тРНК. Для каждой аминокислоты имеется не менее чем по одной специфической тРНК. тРНК связывающие одну и ту же аминокислоту, называют изоакцепторными. Каждый тип клеток в организме отличется своим соотношением изоакцепторных тРНК.
Матричная (информационная)
Матричная РНК содержит генетическую информацию о последовательности аминокислот для основных ферментов и других белков, т.е. служит матицей для биосинтеза полипептидных цепей. На долю мРНК в клетке приходится 5% от общего количества РНК. В отличий от рРНК и тРНК,мРНК гетерогенна по размерам,её молекулярная масса находится в пределах от 25 10 3 до 1 10 6 ; мРНК характеризуется широким диапазоном констант седиментации (6-25S). Наличие в клетке цепи мРНК переменной длинны отражает разнообразие молекулярных масс белков, синтез которых они обеспечивают.
По своему нуклеотидному составу мРНК соответствует ДНК из той же клетки,т.е. является комплементарной к одной из цепи ДНК. В последовательности нуклеотидов (первичная структура) мРНК заложена информация не только о структуре белка, но и о вторичной структуре самих молекул мРНК. Вторичная структура мРНК формируется за счет взаимокомплементарных последовательностей, содержание которых у РНК различного происхождения сходно и состовляет от 40 до 50%. Значительное количество спаренных участков может образовываться в 3 / и 5 / -зонах мРНК.
Анализ 5 / -концов областей 18s рРНК показал,что в них имеются взаимокомплементарные последовательности.
Третичная структура мРНК формируется главным образом за счет водородных связей, гидрофобного взаимодействия, геометрического и стерического ограничения, электрических сил.
Матричная РНК представляет собой метаболически активную и относительно не стабильную, короткоживущую форму. Так, мРНК микроорганизмов характеризуется бысрым обновлением, ивремя жизни её состовляет несколько минут. Вместе с тем для организмов, клетки которых содержат истинные ограниченые мембраной ядра, продолжительность жизни мРНК может достигать многих часов и даже несколько дней.
Стабильность мРНК может определяться различного рода модификациями её молекулы. Так, обнаружено, что 5 / -концевая последовательность мРНК вирусов и эукариот метилирована,или «заблокирована». Первым нуклеотидом в 5 / -терминальной структуре кэпа является 7-метилгуанин, который связан со следующим нуклеотидом 5 / -5 / -пирофосфатной связью. Второй нуклеотид метилирован по C-2 / -рибозного остатка, а в третьем нуклеотиде метильной группы может и не быть.
Ещё одной способностью мРНК является то, что на 3 / -концах многих молекул мРНК эукариотических клеток имеются относительно длинные последовательности адениловых нуклеотидов, которые присоединяются к молекулам мРНК с помощью специальных ферентов уже после завершения синтеза. Реакция протекает в клеточном ядре и цитоплазме.
На 3 / - и 5 / - концах мРНК модифицируемые последовательности составляют около 25% от общей длины молекулы. Считают, что 5 / – кэпы и 3 / -поли-А – последовательности необходимы либо для стабилизации мРНК, предохраняющей её от действия нуклеаз, либо для регулирования процесса трансляции.
РНК-интерференция
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов.
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов.
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5" и 3" нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте - особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
Как и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на:
«минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
двухцепоченые вирусы.
Вироиды - другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина.
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома - ретротранспозоны.

